摘要
Paraplectograptus属包括少数具有相同发育的主要鞘和套表的雷青石种。在试管的正面形成了一个完整的阴道-处女-nema发育和连接列表,而在背面的唯一的鞘列表是侧孔列表,通过套管的胸膜列表连接。输卵管的正面和反面由腹侧的鞘唇连接。许多早期黄脉石的反面的特征之字形鞘表在本属中不存在。在试管的正面和反面,网状安科套表的发育和程度是变化很大的,范围从完全缺乏到密集。Pseudoplectograptus属被认为是Paraplectograptus的初级同义词。Pseudoplectograptus sagenus被转移到sagenograpoides属。其模式种“arctos Sagenograptoides”仅从其伴生的Sagenograptoides sagenus的小的、未成熟的或残缺的标本中被发现。这个物种在腹侧鞘壁上有一个特征性的网状结构,在Paraplectograptus属中没有发现,但在其他Cyrtograptus lundgreni生物带中存在。
介绍
对志留系雷红石笔石属及其管状结构、演化模式和相互关系的认识还存在许多问题。特别是Telychian比大多数其他生物地层层段获得的信息更少,在这些层段中可以发现雷青石(见Maletz 2022,图8)。一些早期分类群,Petalolithinae,提供了重要的信息,显示了安科拉伞发育的各个方面,但可能代表了一般雷青石演化的侧线。它们通常显示完整保存的鞘腹和(成对)孔洞棘,这在任何青石科中都找不到。这些刺可能并不表明与雷青石科的支套发育有系统发育上的联系。早期雷射石属Aeroretiolites和Eorograptus似乎远离这一早期种群,因此它们的结构难以与Petalolithinae的结构进行比较。在Hercograptus属中可以发现它们的螺旋式伞形发育,这是假脉络石的特征,并且可以追溯到pseudoorthograptus obuti或密切相关的petalolithine分类群(见Bates和Kirk 1992)。然而,较年轻的retiolite和Stomatograptus中的ancora伞在其网状发育上有很大的不同,缺乏独特的螺旋结构(见Bates and Kirk 1997)。如果没有少量的化学分离的早期雷顺石,我们对进化历史的理解将局限于伪雷顺石、雷顺石和口纹石这三个强壮的分类群。这些已经显示出相当大的管状分化,其伞形和管状套管的发育表明了强烈的多样化。在系统发育上很难将这些分类群与较年轻的分类群(从Wenlock晚期到Ludlow)联系起来,因为它们的侧索发育较简单。
近年来,关于Paraplectograptus和Pseudoplectograptus及其与粗壮的Retiolites属和Stomatograptus属的潜在进化关系,发表了相当不同的解释。楞次et al。(2018)和Maletz(2022, 2023)认为Paraplectograptus Pseudoplectograptus是同义的,因为它们不具有明确的建筑字符显示足够的分离包括物种,但Kozłowska-Dawidziuk(1995、2004)和贝茨et al。(2023年,p . 52)分离属主要基于他们tubarium维度(“Pseudoplectograptus更健壮,更广泛的rhabdosome而Paraplectograptus苗条,平行的)。
Bates等人(2023)支持长期以来的观点,即从较低的Telychian (llanddovery) Spirograptus turricatus生物带到较低的Homerian (Wenlock) Cyrtograptus lundgreni生物带,存在一个范围很广的Paraplectograptus属(另见Lenz和Melchin 1997,图4;Kozłowska-Dawidziuk 2004,图1;Melchin et al. 2017,图6),显然主要基于未发表的信息和没有标本插图的距离图(参见Lenz and Melchin 1987;Melchin et al. 2017)。Maletz(2022)指出,从Sheinwoodian (Wenlock) Monograptus riccartonensis生物带到Cyrtograptus lundgreni生物带,该属的范围更为有限,拒绝了Melchin等人(2017)的鲜为人知的Paraplectograptus sp.作为Paraplectograptus属的成员。该物种缺乏Paraplectograptus表上的绒状纹饰,具有类似Rotaretiolites的简单anora (Maletz, 2022, p. 11)。
来自离心Cyrtograptus centrifugus Biozone的一个标本最初被鉴定为Paraplectograptus praemacilentus (Štorch 1994, pl. 3,图4),但随后被列为Paraplectograptus cf. eiseli (Štorch 2023, p. 36)。Štorch(2023,图16)显示了该物种从Cyrtograptus insect生物区的上部到Cyrtograptus murchisoni生物区的顶部的范围,其中只有一个标本被展示过。该标本与旁颈龙有明显的相似之处,但不可能进行明确的鉴定。如果这个标本代表Paraplectograptus,那么该属的起源将比本文其他地方提出的要早得多,填补了Maletz(2022)讨论的Telychian到早期Sheinwoodian之间的部分空白。
Bates et al.(2023)完全基于Paraplectograptus可能的早期记录来解释Retiolitinae的早期进化以及Retiolites属和Stomatograptus属的系统发育联系。Kozłowska-Dawidziuk(1995)讨论了Paraplectograptus谱系,当时合并了Paraplectograptus和Pseudoplectograptus属。要了解这两个属的结构细节以及它们与早期雷亚纪的系统发育关系,似乎需要一个新的观点。根据Maletz(2022,图8)的解释,Paraplectograptus必须被视为地层学上相当年轻的雷石系分类群,首先出现在sheinwood dian中部(Wenlock)。雷石属和口纹属起源于早期的特利奇亚(Llandovery),因此至少要早600万年。因此,在我们的记录中有相当长的一段时间,在这段时间里,我们基本上找不到关于复线构造及其演化的资料。由于该组的快速进化,这一差距阻碍了更好地理解雷青石科的管状结构和该组的进化。
Paraplectograptus建设
Paraplectograptus的管状结构是基于许多物种的化学分离材料,因为页岩材料的解释困难且容易导致误解。随着时间的推移,对单个标本的解释已经发生了很大的变化,并且已经添加了可能或可能无法在属中识别的修改。例如,Paraplectograptus (Bates et al. 2023,图2)发现了一个鼻环,但无论是物种的身份还是材料的确切来源(参见Cape Phillips Formation, upper landovery)都无法证实。为了识别属和种,需要有完整标本的插图。标本属Paraplectograptus sp.,来自3号地点附近的一个“孤立结节”;年龄:可能是Llandovery晚期或Wenlock早期’(Lenz 1993,对pl. 13的解释)表明一个类似于Paraplectograptus的分类群,其中保存了具有四个纵向杆的prosicula。
Kozłowska-Dawidziuk (1998, pl. 1,图1)展示了在Zawada钻孔中发现的一个单一标本,即Paraplectograptus eiseli,并保存了一个prosicula。倾斜,扁平的保存使得很难重建管状体的形状和发展,并且Paraplectograptus的分配仍然是暂定的。该标本清楚地代表了一种很少有珊瑚和珊瑚袖表的物种。在插图中无法辨认标本是否有表上的脓疱状纹饰。
之前的解释:
一些相互矛盾的重建已经在文献中提供了副掌。一般来说,这些重建只提供属名,即使它们通常是基于不同物种的特定标本。因此,Lenz等人(2018,图10.4)(图1)对Paraplectograptus的最新重建是基于Lenz(1993,图14,图5)的标本,该标本被确定为Paraplectograptus praemacilentus(图1a, b),该物种后来被称为Pseudoplectograptus (cf.Kozłowska-Dawidziuk 1995;Lenz et al. 2012;Bates et al. 2023)。该标本来自Cape Phillips, CP850 (244-250m;cytograptus lundgreni-睾酮生物区;与Paraplectograptus eiseli, Sokolovograptus textor和其他相关:Lenz 1993, p. 26)。随着时间的推移,重建被修改,现在该标本存在三个明显不同的重建(图1c-e)。Lenz等人(2018)和Bates等人(2023)甚至添加了在标本中无法识别的肩关节环的迹象。从正面来看,解释显示了变化(见图1c左侧的图11;Bates et al. 2005)到反向视图(图1d, e:右侧的ca11;Lenz et al. 2018;Bates et al. 2023)。作为反向视图的解释更有可能,因为试管正面的第一个连接表(th21的基底)向右侧生长(见Maletz 2022,图1E, F:红脉石)。
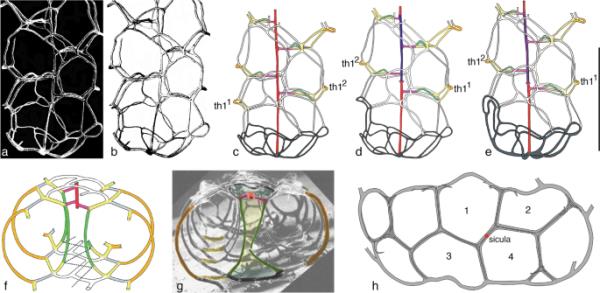
以前的副颈龙重建。a-e Paraplectograptus areticulatus (Kozłowska-Dawidziuk 1995),来自加拿大北极地区的GSC 104044 (a-b)和基于该标本的重建(c-e)。a-b来自Lenz (1993, pl. 14,图5:Paraplectograptus praemacilentus),扫描电镜照片(a)和倒置版本(b)。c Bates等人(2005,图5),解释为正视图中的Pseudoplectograptus。d Lenz等人(2018,图10.4),在反向视图中被解释为附肢。e Bates等人(2023,图1A),被解释为Pseudoplectograptus sp.。标本中未见突起环,但重建图(d-e)显示。比例尺为1mm . f Bates et al.(2023,图1D;根据Lenz, 1993, pl. 14,图3:后来通过Kozłowska-Dawidziuk 1995确定为Pseudoplectograptus areticulatus)。g细纹旁截(Kozłowska-Dawidziuk, 1995),全型,横截面(Maletz 2022,图2C;h Paraplectograptus eiseli (Manck, 1918),伞从外面露出接缝,基于Bates et al.(2023,图3),ZPAL G.47/6;同一标本可能在Bates et al.(2023,图5E)中被描述为Paraplectograptus sp.。所有插图中字符的颜色编码见Maletz(2022,图4)。
Bates等人(2023,图1D)(图1f, g)提到的Paraplectograptus的横截面最初是Lenz和Melchin (1987, pl. 3,图5:Paraplectograptus praemacilentus)的扫描电子显微镜照片,也提供了标本的侧面视图(Lenz和Melchin 1987, pl. 3,图2)Kozłowska-Dawidziuk (1995, p. 290)确定该标本是她的新种Pseudoplectograptus areticulatus的原型。Maletz(2022,图2C)以横截面为例(图1g),确定该物种为Paraplectograptus areticulatus。他的解释与Bates et al.(2023,图1D)提供的解释几乎相同,但后者(图1f)表明,理论上的之字形序列与雷长石的反向之字形序列是同源的,这表明Maletz(2022)的横向序列可能包括部分位于管状体背面的横向孔状序列。
新解释:
由于Paraplectograptus eiseli的模式标本是一种扁平的、保存较差的页岩标本(照片来自Lenz et al. 2012,图7.1),其细节仅部分可用,很难与化学分离的材料进行比较。因此,将化学分离的材料鉴定为Paraplectograptus eiseli可能是值得怀疑的,并且可能不能提供一个合理的例子来鉴定该物种。因此,本文基于Paraplectograptus hermanni Maletz, 2023a,一种与Paraplectograptus eiseli非常相似的物种,但基于化学分离的三维保存材料,对Paraplectograptus属的管状结构提供了新的解释(图2a-c)。
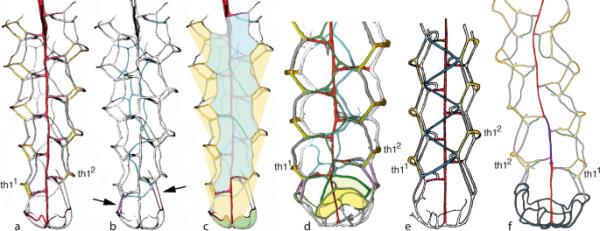
Paraplectograptus解释。a-c Paraplectograptus hermanni Maletz, 2023a,正面全息图的解释。a解释显示阴道-阴道-神经复合体,侧鞘表和胸膜表(袖表)以及彩色鞘唇。b同一样品显示蓝色的反套线。c解释显示鞘膜(黄色),正面伞套膜(浅蓝色)和可能的伞延伸(绿色)。d Paraplectograptus areticulatus (Kozłowska-Dawidziuk, 1995),正视图(基于Palmer and Rickards 1991, pl. 66)。e Paraplectograptus eiseli,解释显示蓝色的反向顶骨表(基于Kozłowska-Dawidziuk 1995,图8C)。f副颌,倒置视图,标本仅显示轻微的侧卧袖表(来自Bates et al. 2023,图1B)。不按比例的标本
hermanni旁膈肌在正面显示了典型的鞘和侧卧袖表发育(图2a)。在这里,virgella-virga-nema复合体与水平连接链相连,与侧孔链和胸膜链的锯齿线相连,形成了输卵管的边缘。正面,因此包括鞘表和胸膜表作为唯一的袖表。在反面,胸膜套管表和侧孔表是仅存的表,从输卵管的轮廓。缺少像在Retiolites和Stomatograptus中看到的锯齿状鞘表(见Maletz 2022,图1E, F)。鞘唇在腹侧连接正反两面。在背面可以看到次要的安科拉套筒列表或网状结构(图2b:蓝色)。在正面的套膜上很少发现次要套表(图2c:绿色)。
anora套筒与anora伞通过四列(紫色列;图2b中箭头所示,此处解释为初始胸膜表。这些列表似乎将ancora伞缘与侧面孔和连接列表的连接处连接起来,并与背面的侧面孔列表连接起来。这些最初的胸膜表可能勾勒出腹侧近侧孔的侧面,正如Bates等人(2023,图4)对Paraplectograptus eiseli提出的那样。因此,不可能判断该分类单元是否发育了近侧孔。因此,在解释中,安科拉袖膜被指示从第一个鞘对的连接列表开始(图2c),但可能起源于更早的位置。在网状旁肌可见明显的外侧近端孔(图2),但未到达胸膜表。
伞的轮廓很薄,很容易折断,因此保存不完整。因此,它的发展很难确定。Bates et al.(2023,图3)绘制了可能属于Paraplectograptus eiseli的标本的伞形图,有9个不同大小的网格(图1)。标本编号(ZPAL47/6,存根313)与Bates等人(2023,图5F)中未显示伞的片段相同。关于Paraplectograptus的伞状结构,没有进一步的信息。
在Paraplectograptus areticulatus中,一个有更广泛的伞套列表的物种,伞套可以解释为延伸到伞的边缘,并且发育包括近侧孔(2d)。相对规则的锯齿状表,其正面和反面与侧孔表和胸膜表的连接处有较短的水平连接,可识别为外侧有接缝的ancora袖表(顶板表)。因此,在背侧的锯齿状鞘表不像在红脉石和口纹石中那样存在。横表的确切发展在拟声部尚不清楚。
安科拉袖表密度:
在不同种类的Paraplectograptus中,侧耳袖表的发育和密度可能变化很大,并且难以区分网状表和网状表(Maletz 2022,第7页)。侧耳袖的胸膜表可以被识别为网状表,与侧孔表一起形成侧耳袖膜的边缘。鉴定侧面之字形顶骨表为克拉石是更有问题的。它们可能非常有规律地发育,如Paraplectograptus areticulatus(图2)。
在背面可以看到比正面更密集的表结构的发展,Paraplectograptus hermanni。试管背面的顶表最初是相当规则的,但在远端不规则性增加(图2b)。在正面只有几个列表可以被注意到(图2c)。
网状发育高度不规则,其次填补了某些类的规整之字形顶链结构,形成了类似于类的不规则网状结构。在其他分类群中也可以看到类似的发展(参见Hoffmanigraptus于Kozłowska 2021,图4A, B;Plectograptus in Bates et al. 2006)。在sagenograpoides sagenus(图3b:蓝色列表)中没有明显的顶表发育,在anora sleeve上发现密集的不规则网状列表。
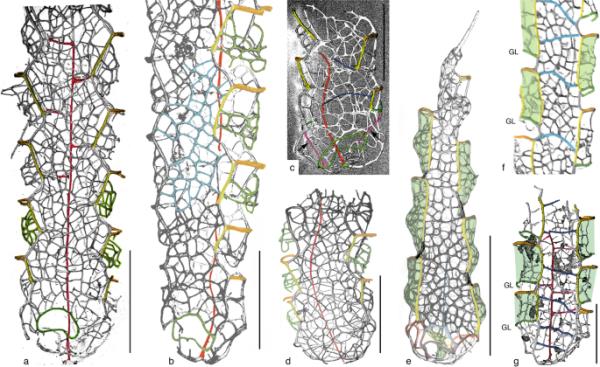
网肌和腹膜壁。a-d sagenograpides sagenus (Lenz, 1993)。a全型图,GSC 104048 (Lenz 1993后,图15,图9)。b成熟管状图,GSC 126924 (Lenz和Kozłowska 2006后,图5.4)。c Sagenograptoides arctos, GSC 119775,正视图(在Lenz和Kozłowska-Dawidziuk 2001, pl. 8之后,图1)。d Sagenograptoides arctos, GSC 99179的全型(在Lenz和Kozłowska-Dawidziuk 2001, pl. 8.5之后)。e Eisenackograptus eisenacki (Obut and Sobolevskaya, 1965), GSC 99146, Arctic Canada (after Lenz 1993, pl. 9.9)。f ? eisenacklograptus eisenacki (Obut and Sobolevskaya, 1965),较长标本的一部分(Kozłowska-Dawidziuk 1990后,图25,图1)。g Virgellograptus perrarus Kozłowska 2015,完整版(Kozłowska 2015后,图1E)。腹侧鞘壁和/或网在这些绿色。比例尺为1mm
Paraplectograptus eiseli的网状发育尚不清楚,因为模式标本(图4a)没有显示所有细节。除了Kozłowska-Dawidziuk(1995,图4)和Lenz etal . (2012, pl. 1,图9)中一个不太好的例子外,尚未证明在化学隔离材料中直接连接反面试管侧边的之字形顶板列表的发展(参见bou
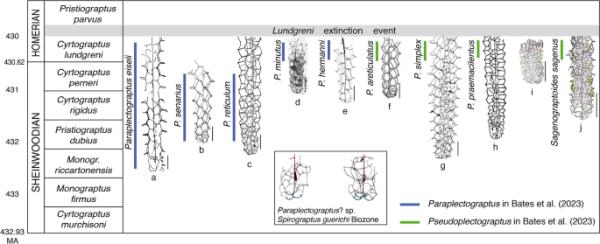
paraplectograpus和sagenograpoides的生物地层学。a Paraplectograptus eiseli(源自Blumenstengel et al. 2006, pl. 1.1)。b senarius Paraplectograptus(根据Lenz et al. 2012,图7.3)。c旁摄网(根据Lenz et al. 2012,图7.4)。d Paraplectograptus minutus(来自Rickards等人,1995年,图20A)。e hermanni副截肢(根据Maletz 2023a,图3G)。f paraectograptus areticulatus(继Palmer and Rickards 1991, pp . 66)。g单纯旁摄图(继Lenz et al. 2012, pl. 2.1之后)。h praemacilentus Paraplectograptus praemacilentus(1952年bou
鞘形:
由于只有少数标本保留了岩鞘壁,故很难观察到岩鞘的真实形态。对Paraplectograptus鞘壁的最佳解释是基于Nancy Kirk准备的模型(见Bates et al. 2023,图7B: as Pseudoplectograptus),因为该属保存的鞘壁膜是未知的。它显示出高度倾斜的腹侧鞘壁和强烈倾斜的孔,没有胫窝或垂直向内倾斜的鞘壁,在许多lundgreni灭绝后的分类群中发现(cf. Kozłowska et al. 2019;Maletz 2023b: Holoretiolites, Neogothograptus)。这种发育(见图2c)与雷铁石(Maletz, 2022,图1E-G)、假雷铁石(Maletz, 2022,图3B)甚至Hercograptus (Melchin, 1999)相似,但此处的腹侧鞘壁倾斜更陡。在sagenograpoides中也可以看到高度倾斜的腹侧鞘壁,部分被网状膜覆盖(图3 -d:绿色)。下Homerian Cyrtograptus lundgreni生物带中lundgreni灭绝前的其他属显示出明显的膝窝和垂直鞘壁(参见Virgellograptus, Cometograptus, Gothograptus, Semigothograptus)。
网状腹膜壁:
在早期脉络膜,侧脉套壁常显示明显的网状链表,但鞘壁上不存在网状链表。鞘壁上的网状网状结构(图3),有时甚至在鞘骨上也是Gothograptus属的特征(参见Kozłowska等人,2019),并且可以在从Cyrtograptus lundgreni生物带开始的其他雷射线中发现(见Maletz 2022,图8:Cometograptus, sagenograpides, Semigothograptus, Virgellograptus)。这种构造的原因和起源都不清楚。在腹侧鞘壁上有网状结构的最古老的分类群包括Eisenackograptus(图3e),其腹侧鞘壁缺乏膝状结构,以及鲜为人知的Virgellograptus(见Kozłowska 2015,图2A),其膝状结构较强(图3g中的GL)。这两个属均首次发现于沈伍德晚期(志留纪文洛克)的Cyrtograptus perneri生物带。
Eisenackograptus在其鞘形态上与sagenograpoides相似,但有一个有限的输卵管,相对较少的输卵管对和一个在阑尾处狭窄的输卵管(参见Kozłowska-Dawidziuk 1990;楞兹,1993)。Eisenackograptus eisenacki (Obut and Sobolevskaya, 1965)具有起伏的网状腹侧鞘壁,至少在第一个鞘对(Maletz 2022,图3C)中可以深入到输卵管(图3C),并且没有膝窝(图3e)。来自波兰Cyrtograptus rigidus Biozone的长标本(Kozłowska-Dawidziuk, 1990, pl. 25)显示出一个独特的没有骨折网的属表(图3f), Kozłowska-Dawidziuk(1995,图9A)中一个标本的重建也是如此。Lenz (1993, pl. 8)和Lenz和Melchin (1987, pl. 3)所描述的一些较长的片段缺少膝窝,并且具有典型eisenacklograptus eisenacki的起伏的腹侧鞘壁。目前尚不清楚,这是否可以被视为单一物种的种内变异,或者这个名称是否涵盖了不止一个物种。
假定的年轻脉络纪的膝骨表与早期笔本类的膝骨表的同源性需要澄清。这些脉冲线系的膝上壁显示网状,而膝表以下没有网状(腱束下壁),也不清楚是否有鞘壁,以及膝表是否可以在结构上(而不是进化上)与气候系的膝带同源,或者代表一个独立发展的特征。
表的表面装饰:
所有的Paraplectograptus物种都具有较年轻的地层学年代学的特征(参见Lenz和Melchin 1987: Plectograptinae;Lenz et al. 2018)。Štorch(1994,2023)鉴定为Paraplectograptus cf. eiseli的材料中是否存在脓疱尚不清楚。根据Maletz(2022,图8)的研究,表面有绒状纹饰的最古老的雷脉石类群属于Giganteograptus Lenz和Kozłowska属(2007)和Sokolovograptus Obut和Zaslavskaya(1974)。
在所有已知的物种中,正面和反面anora袖子的接缝列表在外部都可以找到足够详细的接缝(参见Palmer和Rickards, 1991, pl. 66: Paraplectograptus areticulatus)。在安科拉伞状表的外部也存在接缝(见Bates et al. 2023,图3)。在较年轻的(Wenlock晚期至Ludlow)脉线中,安科拉袖状表在内部缝合(Lenz et al. 2018;Maletz 2022, 2023a)。到目前为止,这两种类型之间的转变还没有被追踪到,但许多晚期的Sheinwoodian物种显示出两者的一些证据(参见Cometograptus: Kozłowska-Dawidziuk 2001)。
Paraplectograptus吗?Melchin et al. sp.(2017)来自Spirograptus guerichi Biozone(基底Telychian, llanddovery),与其他Paraplectograptus不同,Spirograptus的表面纹饰为弱的纵向条纹或光滑的表面。作者指出只有0.4-0.49毫米长,但这显然是保存完好的肩胛骨的长度。现有的标本非常小且不成熟(图4),它们不太可能被纳入Paraplectograptus,因为它们只携带一个明显简单的伞,其列表代表单个鞘对。输卵管远端发育尚不清楚。
胎管长度:
Kozłowska-Dawidziuk(1998,2004)认为脉络膜中脉络膜的长度是理解进化支内进化变化的重要特征。Kozłowska-Dawidziuk (1998, p. 206)提出了一种可能的多种起源,即现在的retiololititiae (Retiolitinae)是基于椎骨的长度。在Paraplectograptus属(Kozłowska-Dawidziuk 2004,图3)中,茎柄的长度显然在1.0到1.5毫米之间变化,但Bates等人(2023,p. 52)的长度为0.9-1.15毫米。镰柄长度在大多数物种中尚未得到证实,因此,不能作进一步的评论。如果副颈龙的标本?Melchin等人(2017)的sp. of Melchin等人(2017)不包括在Paraplectograptus属中,由于没有发表显示prosicula保存的进一步标本,该属的prosicula长度也不为人所知。Kozłowska-Dawidziuk(1995,图4)和Kozłowska-Dawidziuk (1998, pl. 1.1)表明,在Wenlock(? ?)下游地区发现了保存有肩骨的Paraplectograptus eiseli标本。Zawada-1钻芯的Cyrtograptus离心机(Biozone)。然而,这种物质的身份尚不清楚。
星形发生和种内变异
星形发育是笔石分类学中一个经常被忽视的特征,但在分类学上却很重要。Maletz (2023b)讨论了Holoretiolites属和Neogothograptus属的一些物种的星形发育变化,并指出了鉴定幼年标本的问题。除非从样本中恢复了生长序列,否则少于4个卵泡的副栉龙的幼年标本不能归为某一物种。因此,举例来说,Paraplectograptus?来自加拿大北极地区guerichi Spirograptus Biozone(图4:方框)的Melchin等人(2017)在物种水平上无法识别(见Maletz 2022,第11页的注释)。
Paraplectograptus和sagenograpoides的星形发育变化和变异不仅限于标本的最终大小,还包括它们的网状结构密度。哈哥łowska-Dawidziuk (1995;图15C)示出Sagenograptoides sagenus标本,显示出一个有限的管状体。然而,标本远端可能不完整,但它清楚地显示远端网状网密度的减少。此外,Lenz(1993)的标本显示,该物种的网密度远端下降。这种发展可以在次级网存在的Paraplectograptus属的所有物种中看到。在所有的分类群中,网状结构都是最近加入管状结构的,而且大部分网状结构都是在群体形成的相当晚的阶段形成的。
hermanni Paraplectograptus的线虫生长受限(图4e),但大多数物种的菌落最终长度未知。在Paraplectograptus属的任何其他物种中都没有发现线虫,在Sagenograptoides中也没有发现。Manda等人(2019,图10F)展示了一个至少有20对鞘对的Paraplectograptus eiseli标本,这可能是已知最长的Paraplectograptus标本。它显然还在生长,正如远处延伸的nema所显示的那样。其他种类是远端扩大,但没有显示远端锥形的输卵管或指示阑尾表明有限的增长。
种内变异也可能在视网膜线的鉴定中发挥作用(Maletz 2023b),但很少被研究。Paraplectograptus的种内变异从未被探索过,大多数物种都是从很少的标本中描述的。一个很好的例子是Paraplectograptus areticulatus,这个物种只有大约3-4个标本被展示过。该物种看起来是众所周知和常见的,但只有整型标本被一次又一次地说明(参见Lenz和Melchin 1987;Palmer and Rickards 1991;Kozlowska-Dawidziuk 1995;楞次1993;Lenz et al. 2018;Maletz 2022;Bates et al. 2023)。Reichstein(1962)出版的Holoretiolites单纯形(Eisenack, 1935)是唯一的例子的指示不同的种内变异成熟retiolitines标本。
Biostratigraphical分布
Lenz和Melchin(1997,图2)以“Paraplectograptus”的名称描述了一种来自Aeronian (Lituigraptus convolutus Biozone)的早期雷射石的稍微理想化和简化版本,但后来(Melchin et al. 2017, p. 137)将其确定为Eorograptus spirifer。Paraplectograptus吗?来自加拿大北极地区Spirograptus guerichi Biozone的sp. (Melchin et al. 2017, p. 141)被认为是该属中最古老的物种(参见Kozłowska-Dawiziuk 2001),但作者也认为它可能代表一个新属。凤蝶袖表的微纹饰由细小的平行条纹组成,因此不同于凤蝶袖表中的脓疱。在所有已知的标本中,它也有一个完整保存的prosicula。仅鉴定为Paraplectograptus sp.的标本(见Bates et al. 2023,图2C, D)具有增厚的突起边缘,并且缺乏皮质绷带的脓疱表面。不幸的是,这种材料的来源是未知的。另一个来自Llandovery晚期或Wenlock早期的分类群(在一个孤立的结节中发现)具有完整的原体,并显示出ancora袖表的脓疱表面(Lenz 1993, pl. 13,图2)。5-10: Paraplectograptus sp.)与Paraplectograptus没有可比性?p. of Melchin等人(2017)。这些记录都没有证实Paraplectograptus属在sheinwood中期之前的起源。
本文使用最新的数据显示了所有已被接受的Paraplectograptus物种的生物地层范围(图4)。显然最长的生物地层范围是Paraplectograptus eiseli,但该物种的许多鉴定需要验证。Lenz等人(2018,p. 23)指出Paraplectograptus eiseli的选种类型起源于中期telychian Monoclimacis griestoniensis Biozone,尽管Manck(1918)明确指出weterahammer的18区(Monograptus riccartonensis Biozone of Eisel 1899, 1903)在现代生物地层学中大约是中期sheinwoodian。Jaeger(1991,图1)指出在Wetterahammer的Cyrtograptus lundgreni生物带中存在Paraplectograptus eiseli。Lenz et al.(2012)将Paraplectograptus eiseli从Monograptus instrenous - Cyrtograptus kolobus鉴定到加拿大北极的Cyrtograptus perneri生物带,后者与sheinwoodian中部的Monograptus riccartonensis生物带相关(Maletz 2021,图6)。
Lenz et al.(2012)从Monoclimacis fluumendosae到Cyrtograptus perneri生物带中鉴定出了Paraplectograptus reticulum和Paraplectograptus senarius。在成熟标本中,这两个物种的差别主要在于网状结构的密度。所有进一步的物种都只在Cyrtograptus lundgreni生物带中被发现(图4)。数据显示,Paraplectograptus属在生物地层学上局限于sheinwood中期到Homerian基部的区间,并不代表早期的雷石属(参见Bates et al. 2023, p. 46:“最早的雷石属之一”)。在lundgreni灭绝事件中灭绝之前,人们对其系统发育起源和联系以及随后的进化变化知之甚少。粗壮的红脉石和口纹岩具有复杂的链表网,理论上可以简化为在旁纹岩中发现的类型。对安科拉袖网进行相当大的区分和重新排列也是必要的,特别是因为雷石岩没有明确的胸膜表(Maletz 2022,第8页)。
目录
摘要 介绍 Paraplectograptus建设 星形发生和种内变异 Biostratigraphical分布 系统的古生物学 结论 数据可用性 参考文献 致谢 作者信息 道德声明 搜索 导航 #####系统的古生物学
注:笔石目的系统分类遵循最新的论文版本(Lenz et al. 2018;Maletz(协调作者),2023c)。
拉普沃斯亚科(1873年
拟plectograptus bou
诊断:细长,平行侧至远端变宽或有限管;七目伞,边缘不完整;带连接列表的Nema;腹侧由鞘唇,短的外侧孔表和长胸膜表组成;横列存在;安科拉袖表发达到缺乏;安可拉袖表,接缝朝外;带有脓疱的绷带(Maletz(协调作者),2023c,第409页)。
包括种:Retiolites eiseli Manck, 1918;Retiolites tenuis Eisenack, 1951;?布
备注:正如Lenz等人(2018,p. 23)所做的那样,Pseudoplectograptus属与Paraplectograptus同义,但Bates等人(2023)在没有解释的情况下将两者作为单独的属使用。Pseudoplectograptus的模式种为Plectograptus praemacilentus bou
这里提供了所有已知的Paraplectograptus属物种的注释。区分是基于次级或网状安科套表的存在和密度(参见Maletz 2022,第7页)。在附加特征被认为很重要的情况下,对物种的诊断进行了修订。不提供对鲜为人知的物种和同义词的修订。
Paraplectograptus areticculatus (Kozłowska-Dawidziuk, 1995), ex Pseudoplectograptus
(无花果。1 g;4 f)
诊断:输卵管正反两面呈锯齿状顶表的Paraplectograptus种类;在羽绒服袖子上缺少额外的网状列表。
附注:本种发现于加拿大北极环纹环纹生物带。Kozłowska-Dawidziuk (1995, p. 290)称其存在于螺旋体(Spirograptus turrucatus Biozone)到Gothograptus nassa Biozone,但没有发表这种分布的证据。该物种是通过其通常重新绘制的完整模式和Lenz和Melchin(1987)和Lenz(1993)中很少的额外标本而闻名的。没有关于更广泛的生物地理分布的资料。
(曼克,1918),《复刻》
(无花果。1 h ?;2 f;4)
诊断:细长而长,侧面平行的Paraplectograptus种类,有宽环的开口唇;正面与水平连接表连接到nema;试管正面无顶板;在反面的壁表可能在与胸膜表交界处接触;缺乏次级网状。
注:重建Kozłowska-Dawidziuk(1995,图8C;仅鉴定为Paraplectograptus)是误导性的,因为它显示了反面的顶骨列表,但标本的鞘膜对称显示了正面视图(见图2e)。它似乎是基于bou
Paraplectograptus hemmanni bou
赫曼尼·马莱兹,2023a
(无花果。2 a - c;4 e)
诊断:小(5-7鞘对),远端增宽伴明显线状瘤;背面有不规则网状,正面有少量网状。
备注:该物种的特征是一个相对较短的管状体,有一个明显的,细长的线状体,它被合并到管状体的正面(Maletz 2023a,图4H, L)。反面有一个不规则的网状体,但正面只有几个孤立的列表。在腹侧视图中,在近端可以看到不同的伞侧叶(Maletz 2023a,图4H),并且在输卵管的腹侧视图中,在所有Paraplectograptus物种中都可以识别。这些裂片也是后lundgreni灭绝分类群的特征(参见Maletz 2023a,图6C: Gothograptus nassa)。
Paraplectograptus minutus Rickards等人,1995(图4d)。lundgreni/ Biozone,新南威尔士,澳大利亚。该物种可能被称为Paraplectograptus,但管状体的细节尚不清楚,因为材料在页岩中被扁平的标本保存得很差,通常在斜向中发现。该物种网状结构发育适中。
praemacilentus (bou
(无花果。2 d;4小时)
诊断:健壮,最初远端加宽的paraplectograpus种,正反面有不规则的anora套筒列表;单个近腹定向环表在输卵管腹侧。
备注:该物种基于三个页岩标本,在成熟标本的反面和正面似乎显示出相对致密的网状结构。由于标本呈扁平状保存,模式材料中晶格层和网状层的详细结构尚不确定。许多最初属于该物种的标本已被Kozłowska-Dawidziuk(1995)重新分配给Paraplectograptus areticulatus。Lenz等人(2012,p. 16)将“近腹定向环状链表”称为该物种的主要特征。
Paraplectograptus reticulum Lenz等人,2012(图4c)。该物种描述自加拿大北极的Monoclimacis fluendosae至Cyrtograptus perneri生物带。这只是用两个有点变形的标本来说明的。有证据表明,在背袖的反面有网状,但在正面可能没有。
Paraplectograptus senarius Lenz等人,2012(图4b)。加拿大北极地区,与艾斯利拟龙非常相似。不同明显通过发展之字形袖表背面。明显地,eiseli旁肺的壁表呈锯齿状,与侧孔表和胸膜表直接相连(图2e),而senarius旁肺的壁表具有较短的水平连接表(参见图4b)。
单形拟plectograptus (Kozłowska-Dawidziuk, 1995), ex Pseudoplectograptus(图4)。强健的形态具有上部加宽的管状体。安科拉袖网,可能在正面和反面,但细节没有描述。网状不规则,没有清晰的锯齿状壁表。Cyrtograptus lundgreni/ testosterone testis Biozone或Cyrtograptus radians Biozone,波兰。可能是praemacilentus的同义词。
Paraplectograptus tenuis (Eisenack, 1951), ex Retiolites。Eisenack(1951)从许多幼鱼标本中描述了该物种。整型图(Eisenack 1951, pl. 21,图10)显示了一对钙离子。该材料很可能属于Paraplectograptus eiseli或一个密切相关的分类单元,基于近端有一个伞形伞连接到只有四个胸膜表的伞形套。一个特定物种的鉴定是不可能的。
sagenograpides Lenz和Kozłowska, 2010 (=Sagenograptus Lenz和Kozłowska-Dawidziuk, 2001;Sagenograptus Obut和Sobolevskaya的谐音,1962)
诊断:输卵管平行;伞浅;nema免费;近侧口肾形;腹壁由鞘唇,胸膜列,外侧孔列确定;附于伞缘的11和12上的中腹侧表;在茎的第一对的横向表;腹侧鞘壁和鞘套上不规则网状;腹壁表,内有脓疱和缝;安可拉袖表,接缝朝外;可能会出现接缝朝内的外袖表(Maletz(协调作者),2023c,第411页)。
收录种:Paraplectograptus sagenus Lenz, 1993;Sagenograptus arctos Lenz and Kozłowska-Dawidziuk, 2001 (Sagenograptoides sagenus的同义词);在此)。
备注:该属最初由Lenz和Kozłowska-Dawidziuk(2001)命名为Sagenograptus,但由于该名称已用于Tremadocian anisograptid属(Lenz和Kozłowska, 2010),因此不得不重新命名为Sagenograptoides。其模式种Sagenograptoides arctos (Lenz and Kozłowska-Dawidziuk, 2001)在这里被认为是Sagenograptoides sagenus (Lenz, 1993)的初级同义词,Sagenograptoides sagenus最初被描述为Paraplectograptus sagenus。
sagenograpides sagenus (Lenz, 1993) ex Paraplectograptus
(无花果。3模拟;4 i, j)
备注:Sagenograptoides arctos最初鉴定自加拿大北极地区的Cyrtograptus lundgreni/ testosterone testis Biozone。描述是基于少量断裂的近端,显示最多2毫米的长度,有三对鞘膜和输卵管内明显自由的处女-处女-内膜复合体(图3c, d)。它的另一个特征是在侧卧套上和腹侧鞘膜上有致密的网状结构。Sagenograptoides arctos和Paraplectograptus sagenus都来自SBC 10B和SB90 E41m样本(见Lenz和Kozłowska-Dawidziuk 2001),因此是一起出现的。Paraplectograptus sagenus的标本在侧卧套和腹侧鞘膜上具有致密的网状结构(图3a, b)。两个类群的鞘唇都是自由环,而网状结构在假定的鞘膜上开始较低,不接触鞘唇。网状肌显示向外倾斜的腹侧背侧,如腹旁肌。Sagenograptoides sagenus标本比相关的Sagenograptoides arctos标本长得多(至少有6-7鞘对,5毫米长,8毫米长:Lenz 1993, p. 22)。virgella-virga-nema复合物被固定在试管中,至少从第三个鞘对开始,其外侧连接表是可识别的(图3a),但最初似乎是自由的(参见图3b)。
腹侧鞘壁网的发育在许多晚期Sheinwoodian(如Virgellograptus, Cometograptus, Eisenackograptus, Semigothograptus, Gothograptus)系中很常见,但在早期分类群中却很少发现。这是所有后lundgreni灭绝类群的特征(参见Maletz 2023b: Holoretiolites, Neogothograptus)。具有腹侧鞘网的最古老的分类群似乎是Sokolovograptus(见Lenz和Kozłowska 2006,图4:Sokolovograptus cf. S. parens),但这一特征在该属中的分布是相当多变的,正如Sokolovograptus polonicus的例子及其强烈减少的列表网所表明的那样(Kozłowska-Dawidziuk 1995;Lenz et al. 2012)。Sokolovograptus与sagenograpides及其相关分类群的不同之处在于其内部有缝的ancora sleeve list的存在,目前很难建立更紧密的系统发育关系。
腹侧鞘网的发育似乎将许多晚期sheinwooddian与Homerian和更年轻的神经系联系起来,但这一特征的起源尚不清楚。sagenograpoides与Paraplectograptus的关系目前还无法证实,尽管许多结核特征非常相似。
结论
Paraplectograptus属的物种在生物地层上受到限制,从sheinwoodian中期(Monograptus riccartonensis Biozone)到Homerian早期(Cyrtograptus lundgreni Biozone),大多数物种仅在Cyrtograptus lundgreni Biozone中发现。
Paraplectograptus与早期类群的不同之处在于皮质绷带的绒状纹饰和反向之字形鞘表的缺失。安科拉袖表密度变化很大,从密集到缺乏。
安科拉伞看起来很简单,但细节是未知的,安科拉伞的边缘往往难以识别。它可能会减少,一个明显的边缘可能不会发展。
Paraplectograptus在系统发育上可能与腹侧鞘膜上有网状结构的属有关,如sagenograpides、Eisenackograptus、Virgellograptus和其他属。这些分类群也共享具有脓疱表面和外部接缝的列表。
下载原文档:https://link.springer.com/content/pdf/10.1007/s12549-023-00587-x.pdf为您推荐:
- 2023年7月新游戏发布时间、日期和全部阵容 2024-05-11
- 戴安娜王妃在一封信中谈到了与查尔斯离婚后对未来的“恐惧” 2024-05-11
- 巴基斯坦人总理-土耳其的战略合作达到了友谊的新高度 2024-05-11
- 成千上万的人可能不必要地为养老金纳税 2024-05-11
- 当她祈祷时,她欣喜若狂,看到了她的守护天使谁是罗马的弗朗西斯- 2024-05-11
- 拜登任命杰夫·齐恩茨为下一任幕僚长 2024-05-11


